-40%



AMAZING ANCIENT Fossil Coral Reef Sea Life Mineral Specimen MIOCENE MARYLAND
$ 1.05
- Description
- Size Guide
Description
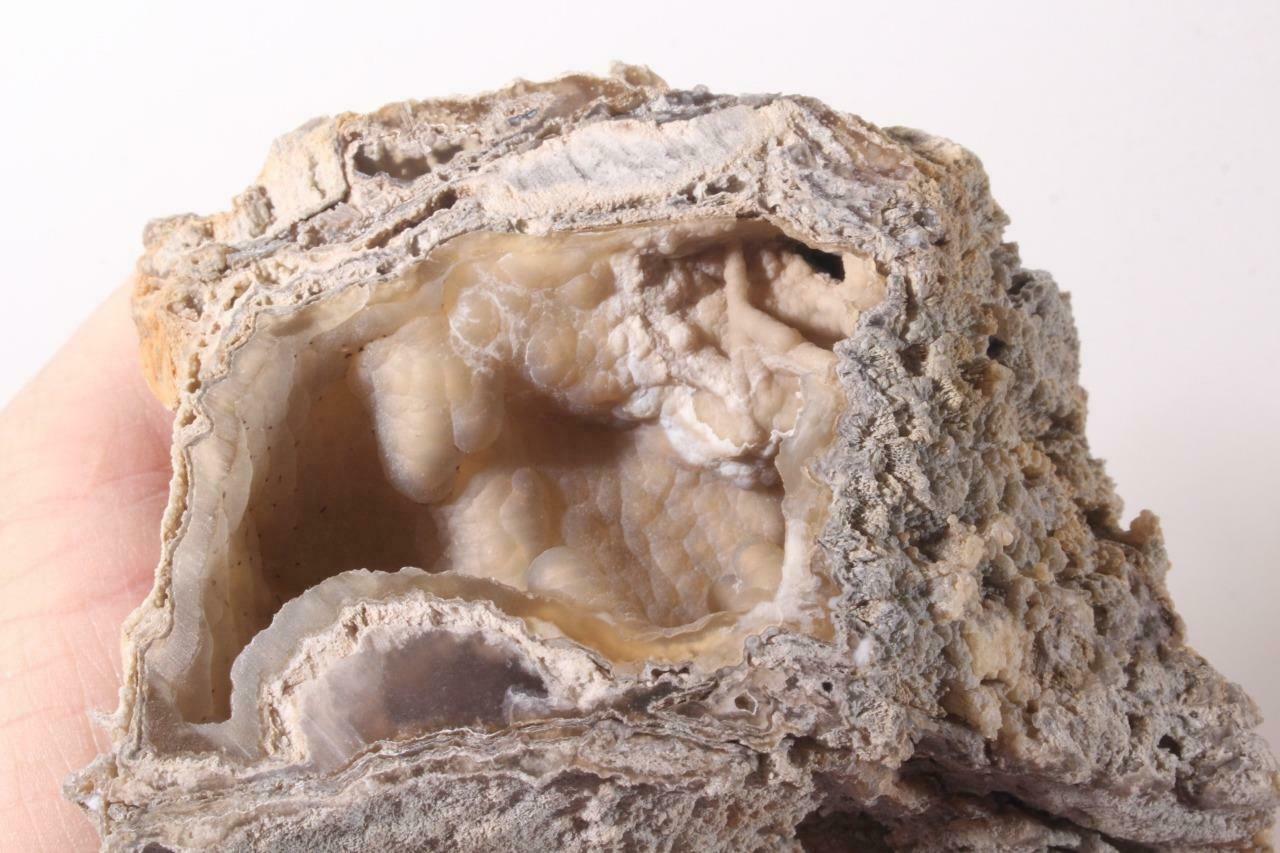
This specimen weighs 8.01 grams. It measures 39 mm x 23 mm x 14 mm.I offer a shipping discount for customers who combine their payments for multiple purchases into one payment!
The discount is regular shipping price for the first item and just 50 cents for each additional item!
To be sure you get your shipping discount just make sure all the items you want to purchase are in your cart.
Auctions you win are added to your cart automatically.
For any "buy it now" items or second chance offers, be sure to click the "add to cart" button, NOT the "buy it now" button.
Once all of your items are in your cart just pay for them from your cart and the combined shipping discount should be applied automatically.
I offer a money back guarantee on every item I sell.
If you are not 100% happy with your purchase just send me a message to let me know
and I will buy back the item for your full purchase price.
Hi there. I am selling this really amazing fossil coral specimen. The species is Astrhelia palmata and it's from the Miocene era, over 5 million years old. I bought it at a gem show in Arizona and it was found in Maryland.It is really amazing, and very detailed. It would be a great idea to have in a classroom for a teacher, or to have as a part of a gem and mineral and fossil collection,It was one of the nicest and most detailed ones that I have ever seen. I have never come across one that looked so complete and perfect. THE DETAIL OF THESE SPECIMENS IS AMAZING!!! It's really interesting and I hope it finds a good home out there. Thanks for visiting my auction! If you have any questions please don't hesitate to ask.
Here is some information about the fossils from Wikipedia. Enjoy, and happy bidding.
Coral
From Wikipedia, the free encyclopedia
For other uses, see Coral (disambiguation).
Coral
Temporal range: Cambrian–Recent
PreЄЄOSDCPTJKPgN
Coral Outcrop Flynn Reef.jpg
A coral outcrop on the Great Barrier Reef, Australia
Scientific classification
Kingdom:
Animalia
Phylum:
Cnidaria
Class:
Anthozoa
Ehrenberg, 1831
Extant subclasses and orders
Octocorallia[1]
Alcyonacea
Helioporacea
Pennatulacea
Hexacorallia[2]
Actiniaria
Antipatharia
Corallimorpharia
Scleractinia
Zoantharia
Ceriantharia[3]
Penicillaria
Spirularia
Corals are marine invertebrates in the class Anthozoa of phylum Cnidaria. They typically live in compact colonies of many identical individual polyps. The group includes the important reef builders that inhabit tropical oceans and secrete calcium carbonate to form a hard skeleton.
A coral "group" is a colony of myriad genetically identical polyps. Each polyp is a sac-like animal typically only a few millimeters in diameter and a few centimeters in length. A set of tentacles surround a central mouth opening. An exoskeleton is excreted near the base. Over many generations, the colony thus creates a large skeleton that is characteristic of the species. Individual heads grow by asexual reproduction of polyps. Corals also breed sexually by spawning: polyps of the same species release gametes simultaneously over a period of one to several nights around a full moon.
Although some corals can catch small fish and plankton using stinging cells on their tentacles, most corals obtain the majority of their energy and nutrients from photosynthetic unicellular dinoflagellates in the genus Symbiodinium that live within their tissues. These are commonly known as zooxanthellae and the corals that contain them are zooxanthellate corals. Such corals require sunlight and grow in clear, shallow water, typically at depths shallower than 60 metres (200 ft). Corals are major contributors to the physical structure of the coral reefs that develop in tropical and subtropical waters, such as the enormous Great Barrier Reef off the coast of Queensland, Australia.
Other corals do not rely on zooxanthellae and can live in much deeper water, with the cold-water genus Lophelia surviving as deep as 3,000 metres (9,800 ft).[4] Some have been found on the Darwin Mounds, north-west of Cape Wrath, Scotland. Corals have also been found as far north as off the coast of Washington State and the Aleutian Islands.
Contents [hide]
1
Taxonomy
2
Anatomy
3
Ecology
3.1
Feeding
3.2
Intracellular symbionts
4
Reproduction
4.1
Sexual
4.2
Asexual
4.3
Colony division
5
Reefs
6
Evolutionary history
7
Status
7.1
Threats
7.2
Protection
8
Relation to humans
8.1
Jewelry
8.2
Medicine
8.3
Construction
8.4
Climate research
8.5
Aquaria
8.6
Aquaculture
9
Gallery
10
References
11
Sources
12
External links
Taxonomy[edit]
Hexacorallia
Actiniaria
Antipatharia
Corallimorpharia
Scleractinia
Zoantharia
Octocorallia
Alcyonacea
Helioporacea
Pennatulacea
Ceriantharia
Penicillaria
Spirularia
Phylogeny of Anthozoa
In his Scala Naturae, Aristotle classified corals as "zoophyta" ("plant-animals"), animals that had characteristics of plants and were therefore hypothetically in between animals and plants. The Persian polymath Al-Biruni (d. 1048) classified sponges and corals as animals, arguing that they respond to touch.[5] Nevertheless, people believed corals to be plants until the eighteenth century, when William Herschel used a microscope to establish that coral had the characteristic thin cell membranes of an animal.[6]
The phylogeny of Anthozoans is not clearly understood and a number of different models have been proposed. Within the Hexacorallia, the sea anemones, coral anemones and stony corals may constitute a monophyletic grouping united by their six-fold symmetry and cnidocyte trait. The Octocorallia appears to be monophyletic, and primitive members of this group may have been stolonate.[7] The cladogram presented here comes from a 2014 study by Stampar et al. which was based on the divergence of mitochondrial DNA within the group and on nuclear markers.[8]
Corals are classified in the class Anthozoa of the phylum Cnidaria. They are divided into three subclasses, Hexacorallia, Octocorallia,[9] and Ceriantharia.[8][10] The Hexacorallia include the stony corals, the sea anemones and the zoanthids. These groups have polyps that generally have 6-fold symmetry. The Octocorallia include blue coral, soft corals, sea pens, and gorgonians (sea fans and sea whips). These groups have polyps with 8-fold symmetry, each polyp having eight tentacles and eight mesenteries. Ceriantharia are the tube-dwelling anemones.[7]
Fire corals are not true corals, being in the order Anthomedusa (sometimes known as Anthoathecata) of the class Hydrozoa.[11]
Anatomy[edit]
Anatomy of a stony coral polyp
Corals are sessile animals in the class Anthozoa and differ from most other cnidarians in not having a medusa stage in their life cycle. The body unit of the animal is a polyp. Most corals are colonial, the initial polyp budding to produce another and the colony gradually developing from this small start. In stony corals, also known as hard corals, the polyps produce a skeleton composed of calcium carbonate to strengthen and protect the organism. This is deposited by the polyps and by the coenosarc, the living tissue that connects them. The polyps sit in cup-shaped depressions in the skeleton known as corallites. Colonies of stony coral are very variable in appearance; a single species may adopt an encrusting, plate-like, bushy, columnar or massive solid structure, the various forms often being linked to different types of habitat, with variations in light level and water movement being significant.[7]
In soft corals, there is no stony skeleton but the tissues are often toughened by the presence of tiny skeletal elements known as sclerites, which are made from calcium carbonate. Soft corals are very variable in form and most are colonial. A few soft corals are stolonate, but the polyps of most are connected by sheets of coenosarc. In some species this is thick and the polyps are deeply embedded. Some soft corals are encrusting or form lobes. Others are tree-like or whip-like and have a central axial skeleton embedded in the tissue matrix. This is composed either of a fibrous protein called gorgonin or of a calcified material. In both stony and soft corals, the polyps can be retracted, with stony corals relying on their hard skeleton and cnidocytes for defence against predators, with soft corals generally relying on chemical defences in the form of toxic substances present in the tissues known as terpenoids.[7]
Montastraea cavernosa polyps with tentacles extended
The polyps of stony corals have six-fold symmetry while those of soft corals have eight. The mouth of each polyp is surrounded by a ring of tentacles. In stony corals these are cylindrical and taper to a point, but in soft corals they are pinnate with side branches known as pinnules. In some tropical species these are reduced to mere stubs and in some they are fused to give a paddle-like appearance.[12] In most corals, the tentacles are retracted by day and spread out at night to catch plankton and other small organisms. Shallow water species of both stony and soft corals can be zooxanthellate, the corals supplementing their plankton diet with the products of photosynthesis produced by these symbionts.[7] The polyps interconnect by a complex and well-developed system of gastrovascular canals, allowing significant sharing of nutrients and symbionts.[13]
Ecology[edit]
Feeding[edit]
Polyps feed on a variety of small organisms, from microscopic zooplankton to small fish. The polyp's tentacles immobilize or kill prey using their nematocysts. These cells carry venom which they rapidly release in response to contact with another organism. A dormant nematocyst discharges in response to nearby prey touching the trigger (cnidocil). A flap (operculum) opens and its stinging apparatus fires the barb into the prey. The venom is injected through the hollow filament to immobilise the prey; the tentacles then manoeuvre the prey to the mouth.[14]
The tentacles then contract to bring the prey into the stomach. Once the prey is digested, the stomach reopens, allowing the elimination of waste products and the beginning of the next hunting cycle. They can scavenge drifting organic molecules and dissolved organic molecules.[15]:24
Intracellular symbionts[edit]
Many corals, as well as other cnidarian groups such as Aiptasia (a sea anemone) form a symbiotic relationship with a class of dinoflagellate algae, zooxanthellae of the genus Symbiodinium.[15]:24 Aiptasia, a familiar pest among coral reef aquarium hobbyists, serves as a valuable model organism in the study of cnidarian-algal symbiosis. Typically, each polyp harbors one species of algae. Via photosynthesis, these provide energy for the coral, and aid in calcification.[16] As much as 30% of the tissue of a polyp may be algal material.[15]:23
The algae benefit from a safe place to live and consume the polyp's carbon dioxide and nitrogenous waste. Due to the strain the algae can put on the polyp, stress on the coral often drives them to eject the algae. Mass ejections are known as coral bleaching, because the algae contribute to coral's brown coloration; other colors, however, are due to host coral pigments, such as green fluorescent proteins (GFPs). Ejection increases the polyp's chance of surviving short-term stress—they can regain algae, possibly of a different species at a later time. If the stressful conditions persist, the polyp eventually dies.[17]
Reproduction[edit]
Corals can be both gonochoristic (unisexual) and hermaphroditic, each of which can reproduce sexually and asexually. Reproduction also allows coral to settle in new areas.
Sexual[edit]
Life cycles of broadcasters and brooders
Corals predominantly reproduce sexually. About 25% of hermatypic corals (stony corals) form single sex (gonochoristic) colonies, while the rest are hermaphroditic.[18]
Broadcasters[edit]
About 75% of all hermatypic corals "broadcast spawn" by releasing gametes—eggs and sperm—into the water to spread offspring. The gametes fuse during fertilization to form a microscopic larva called a planula, typically pink and elliptical in shape. A typical coral colony forms several thousand larvae per year to overcome the odds against formation of a new colony.[19]
A male great star coral, Montastraea cavernosa, releasing sperm into the water.
Synchronous spawning is very typical on the coral reef, and often, even when multiple species are present, all corals spawn on the same night. This synchrony is essential so male and female gametes can meet. Corals rely on environmental cues, varying from species to species, to determine the proper time to release gametes into the water. The cues involve temperature change, lunar cycle, day length, and possibly chemical signalling.[18] Synchronous spawning may form hybrids and is perhaps involved in coral speciation.[20] The immediate cue is most often sunset, which cues the release.[18] The spawning event can be visually dramatic, clouding the usually clear water with gametes.
Brooders[edit]
Brooding species are most often ahermatypic (not reef-building) in areas of high current or wave action. Brooders release only sperm, which is negatively buoyant, sinking on to the waiting egg carriers who harbor unfertilized eggs for weeks. Synchronous spawning events sometimes occurs even with these species.[18] After fertilization, the corals release planula that are ready to settle.[16]
Planulae[edit]
Planula larvae exhibit positive phototaxis, swimming towards light to reach surface waters, where they drift and grow before descending to seek a hard surface to which they can attach and begin a new colony. They also exhibit positive sonotaxis, moving towards sounds that emanate from the reef and away from open water.[21] High failure rates afflict many stages of this process, and even though millions of gametes are released by each colony, few new colonies form. The time from spawning to settling is usually two to three days, but can be up to two months.[22] The larva grows into a polyp and eventually becomes a coral head by asexual budding and growth.
Asexual[edit]
Basal plates (calices) of Orbicella annularis showing multiplication by budding (small central plate) and division (large double plate)
Tabulate coral Aulopora (Devonian) showing initial budding
Within a coral head, the genetically identical polyps reproduce asexually, either by budding (gemmation) or by dividing, whether longitudinally or transversely.
Budding involves splitting a smaller polyp from an adult.[19] As the new polyp grows, it forms its body parts. The distance between the new and adult polyps grows, and with it, the coenosarc (the common body of the colony). Budding can be intratentacular, from its oral discs, producing same-sized polyps within the ring of tentacles, or extratentacular, from its base, producing a smaller polyp.
Division forms two polyps that each become as large as the original. Longitudinal division begins when a polyp broadens and then divides its coelenteron (body), effectively splitting along its length. The mouth divides and new tentacles form. The two polyps thus created then generate their missing body parts and exoskeleton. Transversal division occurs when polyps and the exoskeleton divide transversally into two parts. This means one has the basal disc (bottom) and the other has the oral disc (top); the new polyps must separately generate the missing pieces.
Asexual reproduction offers the benefits of high reproductive rate, delaying senescence, and replacement of dead modules, as well as geographical distribution.[23]
Colony division[edit]
Whole colonies can reproduce asexually, forming two colonies with the same genotype. The possible mechanisms include fission, bailout and fragmentation. Fission occurs in some corals, especially among the family Fungiidae, where the colony splits into two or more colonies during early developmental stages. Bailout occurs when a single polyp abandons the colony and settles on a different substrate to create a new colony. Fragmentation involves individuals broken from the colony during storms or other disruptions. The separated individuals can start new colonies.[24]
Reefs[edit]
Locations of coral reefs around the world
Main article: Coral reef
See also: Coral reef fish and List of reefs
Many corals in the order Scleractinia are hermatypic, meaning that they are involved in building reefs. Most such corals obtain some of their energy from zooxanthellae in the genus Symbiodinium. These are symbiotic photosynthetic dinoflagellates which require sunlight; reef-forming corals are therefore found mainly in shallow water. They secrete calcium carbonate to form hard skeletons that become the framework of the reef. However, not all reef-building corals in shallow water contain zooxanthellae, and some deep water species, living at depths to which light cannot penetrate, form reefs but do not harbour the symbionts.[25]
Staghorn coral (Acropora cervicornis) is an important hermatypic coral from the Caribbean
There are various types of shallow-water coral reef, including fringing reefs, barrier reefs and atolls; most occur in tropical and subtropical seas. They are very slow-growing, adding perhaps one centimetre (0.4 in) in height each year. The Great Barrier Reef is thought to have been laid down about two million years ago. Over time, corals fragment and die, sand and rubble accumulates between the corals, and the shells of clams and other molluscs decay to form a gradually evolving calcium carbonate structure.[26] Coral reefs are extremely diverse marine ecosystems hosting over 4,000 species of fish, massive numbers of cnidarians, molluscs, crustaceans, and many other animals.[27]
Evolutionary history[edit]
Solitary rugose coral (Grewingkia) in three views; Ordovician, southeastern Indiana
Although corals first appeared in the Cambrian period,[28] some 542 million years ago, fossils are extremely rare until the Ordovician period, 100 million years later, when rugose and tabulate corals became widespread. Paleozoic corals often contained numerous endobiotic symbionts.[29][30]
Tabulate corals occur in limestones and calcareous shales of the Ordovician and Silurian periods, and often form low cushions or branching masses of calcite alongside rugose corals. Their numbers began to decline during the middle of the Silurian period, and they became extinct at the end of the Permian period, 250 million years ago.[31]
Rugose or horn corals became dominant by the middle of the Silurian period, and became extinct early in the Triassic period. The rugose corals existed in solitary and colonial forms, and were also composed of calcite.[32]
The scleractinian corals filled the niche vacated by the extinct rugose and tabulate species. Their fossils may be found in small numbers in rocks from the Triassic period, and became common in the Jurassic and later periods.[33] Scleractinian skeletons are composed of a form of calcium carbonate known as aragonite.[34] Although they are geologically younger than the tabulate and rugose corals, the aragonite of their skeletons is less readily preserved, and their fossil record is accordingly less complete.